Turing's diffusion-driven instability is a widely used canonical model for pattern formation in biology. The basic Turing model consists of two coupled diffusion-reaction partial differential equations. Traditionally, the model is posed on a fixed domain. Depending on the kinetic terms and the parameters, the typical resulting patterns are spots, and/or labyrinthine stripes.
However, many stripe patterns observed in biology are not disordered labyrinths, but instead they are aligned along a specific direction. Consider a zebra: the stripes along most of its body are vertical. What mechanisms can cause this kind of robust directionality and alignment?
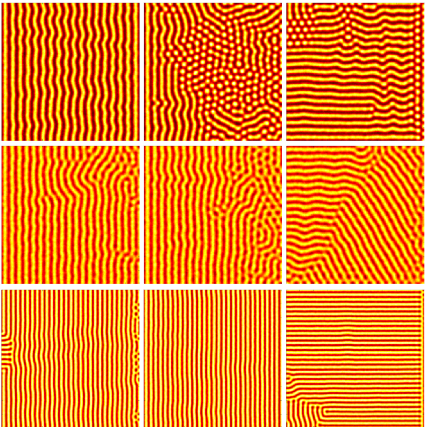
A wave of competency refers to a phenomenon where the region where patterning is possible expands out like a wave, usually due to maturation of the underlying tissue. We show that it is possible to select a preferred alignment and directionality of the stripe patterns by modulating the wave speeds. Moreover, this holds true for multiple different models. In the figure on the left, the columns from left to right corresponds to decreasing wave speeds. We postulate that this mechanism can explain the stripes found in many biological systems, and the wave of competency can be used to control the patterning process, in order to obtain a desired pattern.